
Table of Contents
Ecdysteroid metabolism
Ecdysteroids is a generic term for the arthropod molting hormones, including ecdysone, first isolated from Bombyx pupae in 1954, 20-hydroxyecdysone (20E), the most common molting hormone, and related molecules of which many are found in plants. According to Lafont et al. (2012) ecdysteroids can be defined either by their biological activity (molting hormones) or their chemical structure. They represent a specific family of sterol derivatives that bear common structural features: a cis (5β-H) junction of rings A and B, a 7-ene-6-one chromophore, and a trans (14α-OH or H) junction of rings C and D. Most of them also bear the 3β-OH already present in their sterol precursor, and a set of other hydroxyl groups located both on the steroid nucleus and the side chain. The major insect ecdysteroid is 20E, a 27-carbon (C27) molecule derived from cholesterol, but some insect species contain its C28 or C29 homologs (methyl or ethyl at C24), respectively.
P450 and ecdysteroid biosynthesis
The role of P450 enzymes in ecdysteroid metabolism has been known for many years (review in Lafont et al., 2012) and several CYP genes have clearly been associated with ecdysteroid biosynthesis. These are the CYP2 clan CYP306A1 and CYP307 as well as the mitochondrial clan CYP302A1, CYP314A1 and CYP315A1. The following figure shows 20-hydroxyecdysone with carbon numbering and the currently known sites of P450 metabolism. The CYP2 clan CYP18 is also a P450 involved in ecdysteroid metabolism. In this figure, the exact function of CYP307 is not defined.
These genes are often called by their confusing Drosophila mutant names, Cyp306a1 (phantom, phm), Cyp307a1 (spook, spo), Cyp307a2 (spookier, spok), Cyp302a1 (disembodied, dib), Cyp314a1 (shade, shd), and Cyp315a1 (shadow,sad). They were collectively dubbed Halloween genes, a term that does not appear in the original (1984) publications from the Nüsslein-Volhard lab describing the mutants, and that obscures the relationship with CYP18. They should just be called ecdysteroidogenic P450s for convenience (see also review in Niwa and Niwa, 2014). This denomination is clearer and allows expansion to the P450s involved in ecdysteroid metabolism that remain to be characterized.
There are also genes in the so-called “Halloween” series (e.g. neverland,and non-molting glossy /shroud) that are not P450 genes, and this creates additional confusion, with some authors drawing phylogenies of “Halloween” genes that include P450 and non-P450 sequences.
The reactions in ecdysteroid biosynthesis from sterol precursors are generally considered to be structured in a linear pathway: 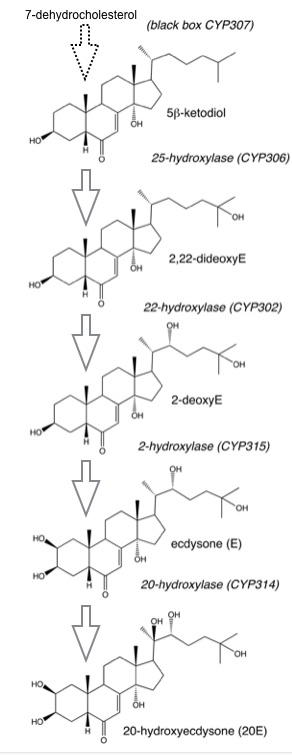
In this pathway as classical depicted, the sterol precursor would be the C27 cholesterol. In some insects, C28 or C29 sterols are used instead, leading to C24-methyl or ethyl-branched ecdysteroids. Furthermore, the product of the black box depicted here is a 3-hydroxysterol, but in some insects it may be a 3-ketosterol, with the 3 keto group reduced during or after ecdysone production. P450 genes currently known to catalyze the steps depicted are mentioned in brackets.
The origin of the P450 genes involved in ecdysteroid biosynthesis is of great evolutionary importance, as one might expect “Ecdysozoa” to be capable of making a molting hormone. However, the presence of ecdysteroids, or indeed of the ecdysteroidogenic P450s, is clearly not a synapomorphy (a shared derived trait that indicates a common ancestry) of “Ecdysozoa”. For instance the well-known nematode model Caenorhabditis elegans has no ecdysteroids and none of the P450s needed to make them.
Deviations from this scheme (see below with ecdysteroids identified in arthropods and their possible origin from a non-linear pathway) are rarely discussed and experimental evidence for the activity or specificity of the enzymes is limited to a few species (see individual steps below). The most common deviation is the lack of C25 hydroxylation, as in spider mites (Grbic et al., 2011) leading to ponasterone A as major ecdysteroid. Barnacles (Thecostraca, Cirripedia) are particularly startling cases of “missing” genes, which highlight the limitations of the “Halloween” gene denomination and the need for further searches of ecdysteroidogenic P450s. These sessile organisms clearly molt, and clearly have 20-hydroxyecdysone and ecdysone (Bebbington and Morgan, 1977). Yet extensive searches of the many TSA and three barnacle genomes failed to recover any trace of CYP302, CYP314, CYP18 or CYP306. Instead, only CYP307 (duplicated) and CYP315 were found. All barnacle CYP315 also had a peculiar insertion between the PERW and Cys pocket motifs. The other CYP2 clan and mitochondrial P450s of barnacles are all paralogs of known CYP families in Crustacea, rather than being just highly divergent CYP18/306 or CYP302 and CYP314.
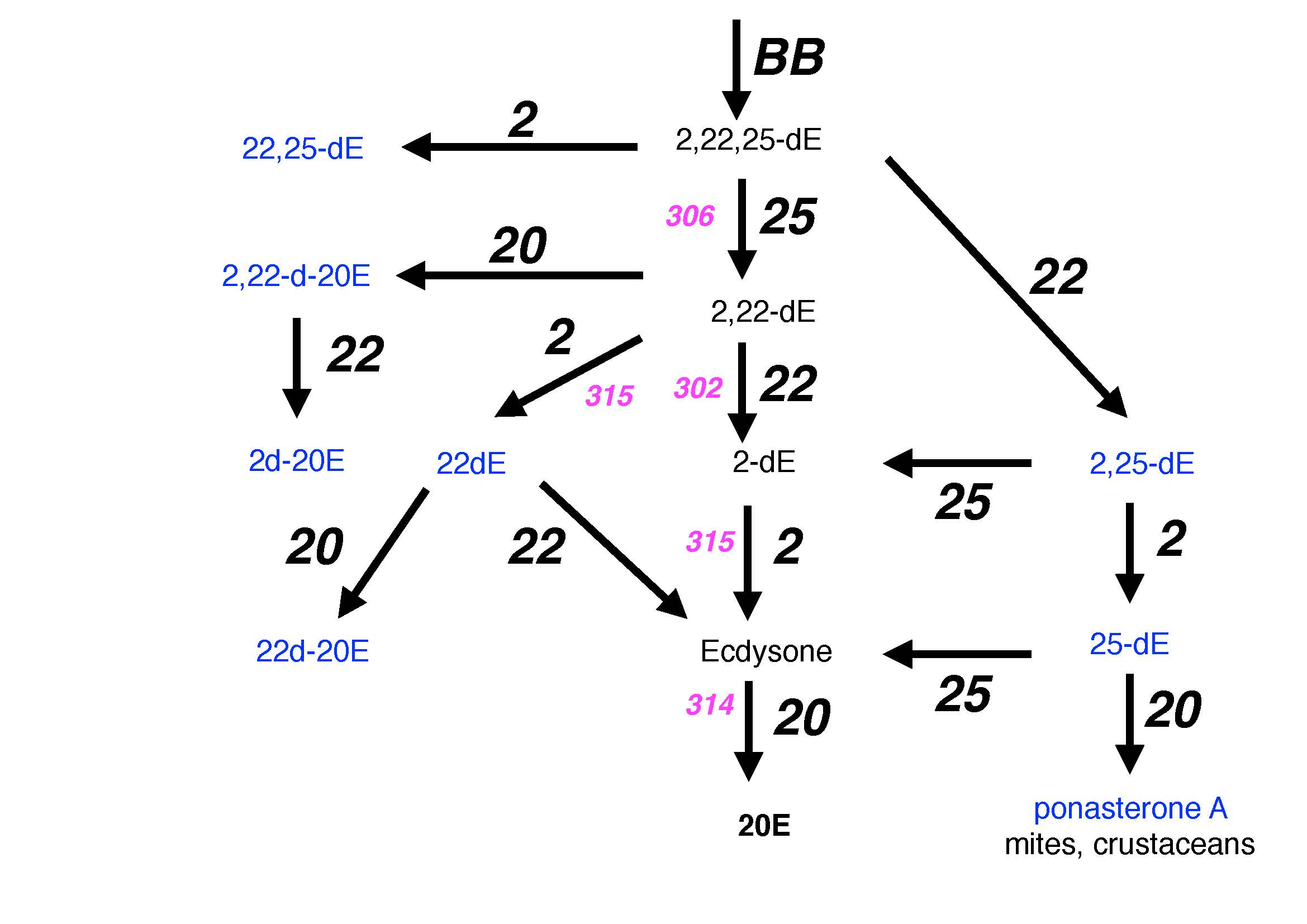
P450 catalyzed steps in ecdysteroid biosynthesis
Early steps, the "black box" and CYP307
CYP306A1 was identified as a C25 hydroxylase in Bombyx mori and in Drosophila. The C25 hydroxylase of Chelicerata has not been identified yet.
CYP302A1 is an ecdysteroid C22 hydroxylase also called disembodied (dib) in Drosophila (Warren et al., 2002).
CYP315A1 is an ecdysteroid C2 hydroxylase also called shadow (sad) in Drosophila (Warren et al., 2002).
CYP314A1 is an ecdysteroid C20 hydroxylase also called shade (shd) in Drosophila (Petryk et al., 2003).
P450 and ecdysteroid catabolism
C26 hydroxylation/oxidation
In insects, CYP18A1 metabolizes ecdysteroids by hydroxylation and further oxidation at C26. This constitutes hormonal inactivation in Drosophila (Guittard et al., 2011).
The C-26 hydroxylation of 20-hydroxyecdysone has been characterized as a typical microsomal P450 activity in an epithelial cell line of Chironomus tentans (Kayser et al., 1997). However, a dual localization (microsomal and mitochondrial) was reported for ecdysteroid 26-hydroxylase in Manduca sexta midgut (Williams et al., 1997, 2000). The reaction has a low Km for 20-hydroxyecdysone (~ 1 μM). The lepidopteran midgut C26-hydroxylase would presumably be encoded by CYP18B1 as in B. mori (Li et al., 2014). Indeed Manduca sexta CYP18B1 has a predicted mitochondrial location by DeepLoc1.0. While this is unusual for a CYP2 clan P450, it would explain the biochemical data (Williams et al., 2000) yet would suggest, in addition, the existence of a microsomal P450 with C26 hydroxylase activity, or a cryptic microsomal targeting sequence in CYP18B1.
The Chironomus tentans cell line metabolizes 20,26-dihydroxyecdysone further to two less polar metabolites produced in a constant 3:1 ratio. This metabolism is NADPH-dependent and inhibited by azole compounds. The two metabolites are diastereomers of a cyclic hemiacetal formed (nonenzymatically) by the reaction of the C-22 hydroxyl group with a C-26 aldehyde (Kayser et al., 2002). The C-26 aldehyde is a presumed intermediate in the conversion of C-26 ecdysteroids to C-26 ecdysonoic acids, a common inactivation product of ecdysteroids (Rees, 1995). It is now clearly established that the P450 conversion of the C-26 hydroxyl to the C-26 carboxylic acid is carried out by the same enzyme that initially hydroxylates 20-hydroxyecdysone (see figure adapted from Guittard et al., 2011).
This enzyme was shown in Drosophila (Guittard et al., 2011) to be encoded by the CYP18A1 gene. Depending on the species, and perhaps tissues, the reaction proceeds to the C26-hydroxylated product, the cyclic hemiacetals, or the 26-carboxylic acid. The Chironomus tentans enzyme metabolizes ecdysone, 20-hydroxyecdysone, but also ponasterone A (25-deoxy-20E)(Kayser et al., 2011). The conversion of ponasterone A to inokosterone (26-hydroxy-ponasterone A) occurs preferentially to the 25R epimer.
Shi et al., 2022 reported a modest O-dealkylation activity of Helicoverpa armigera CYP18A1 towards 7-ethoxycoumarin (7EC) and epoxidation activity towards aldrin, while CYP18B1 was able to dealkylate 7-benzyloxymethoxy resorufin (BOMR).
The expression of CYP18A1 pulses closely after each ecdysteroid peak in Drosophila (Bassett et al., 1997). CYP18A1 null alleles or RNAi knockdown of CYP18A1, as well as ectopic overexpression of CYP18A1 cause severe phenotypes and lethality, thus highlighting the important regulatory role of the enzyme in ecdysteroid inactivation in insects (Rewitz et al., 2010; Guittard et al., 2011).
Side chain cleavage
An early report of ecdysteroid catabolism by side-chain cleavage to poststerone in Bombyx mori (Hikino et al., 1975) has not been followed up in the literature. Despite the analogy to the reaction catalyzed by vertebrate P450scc (CYP11A), i.e. the C-C bond cleavage at a vicinal C-20,C-22 diol, there is currently no information on the enzymology of this reaction in insects. P450scc is a unique enzyme in that it catalyzes not just the C-C bond cleavage, but also the two preceding hydroxylations of cholesterol at C-20 and C-22 (in that order). In the case of arthropods, these two hydroxylations occur independently.